

細菌、藻類及真菌類 (中卷:藻類)

概論
藻類為延陵植物志第一部分的中卷, 為繼細菌類以後的章節, 從植物的自然歷史上觀之, 可說是最關鍵的進化階段, 它們承繼高等細菌成為更高階的植物類群, 而且它們演化出先進的光合作用機制, 為將來出現的苔蘚類、蕨類及種子植物提供了必要的基礎, 同時有些藻類且具備原生動物細胞的特徵, 故外, 它們與動物的起源也有一定關連。
藻類是具有葉綠素、能進行尤合作用、營自養生活的無維管束、無胚的葉狀體植物, 一般生長在水體中。藻類植物有兩個特點: (1)藻體各式各樣,在外形上有時類似高等植物的根、莖、葉的構造, 但在功能上都能進行光合作用、釋放氧氣, 因而實際上藻體就是一個簡單的葉, 也因此藻類植物的藻體統稱為葉狀體; (2)它們的有性生殖器官一般都為單細胞,有的可以是多細胞的, 但缺少一層包圍的營養細胞, 所有細胞都直接參與生殖作用。
藻類植物的起源及分化
藻類植物可以是從原始的光合細菌發展而來的。光合細菌具有細菌綠素, 利用元機的硫化氫作為氫的供應者,
產生了光系統 I 。原始藻類植物, 如藍藻類所具有的葉綠素a, 很可能是由細菌綠素進化而來的。藍藻類利用廣泛存在的水為氫的供應者, 具有光系統II,
通過光合作用產生了氧。隨著藍藻類的產生, 光合細菌類逐漸退居次要地位, 而放氧型的藍藻類則逐漸成為佔優勢的種類, 釋放出來的氧氣逐漸改變了大氣性質,
使整個生物界朝著能量利用效率更高的喜氧生物方向發展。這個方向的進一步發展就產生了具有真核的紅藻類, 同時, 類囊體單條地組成為葉綠體, 但集光色素基本上一樣,
仍以藻膽蛋白為集光色素。藍藻和紅藻的集光色素, 藻膽蛋白, 需用大量能量和物質合成, 是很不經濟的原始類型, 所以只能發展到紅藻類, 形成進化上的一個盲枝。
藻類植物的第二個發展方向是在海洋裏產生含葉綠素a和葉綠素c的雜色藻類。葉綠素c代替了藻膽蛋白, 進一步解決了更有效地利用光能的問題。在開始的時候,
藻膽蛋白仍繼續存在, 如在隱藻類, 但進一步的進化, 效率較低的藻膽蛋白沒有繼續存在的必要而逐漸被淘汰, 所以在比隱藻類較為高級的種類, 如在甲藻類、矽藻類,
除葉綠素a以外, 只有葉綠素c, 而藻膽蛋白消失了。迄今, 海洋仍為含有葉綠素c的種類,
包括甲藻類、金藻類、黃藻類和矽藻類等浮游藻類和褐藻類的底棲藻類而佔據優勢。但這個類群不能離開水體, 仍是一個盲校。
藻類植物的第三發展方向是在海洋較淺處產生綠色植物。它們除了葉綠素a以外, 還產生了葉綠素b。據科學家估計, 葉綠素 a+b系統比之葉綠素a+藻膽蛋白系統,光合作用效率高出了3倍,
也高於葉綠素a+c 系統。這是藻類植物進化的主流。很可能十幾年前發現的原綠藻就是這類植物的祖先。原綠藻植物出現的時間可能與原核的雜色藻類差不多,
但由於某種原因, 可能 與當時的大氣光照條件有關, 雜色藻類大量發展起來而原綠藻卻停留在原始狀態。後來環境條件變為較為適 合於葉綠素b 生物的生長,
從原綠藻植物就產生了真核 的綠藻類。它們不但已產生了葉綠體, 而且已經有了比較其他藻類更加進步的光合器, 即具有基粒的葉綠體。
就是這類植物終於登陸, 進一步演化為苔蘚植物、蕨類植物及種子植物。幾億年前地球大氣的含氧量已達到現在大氣的百分之十, 形成了臭氧遮罩層,
阻擋了殺傷生物的紫外線, 使陸地具備了生命生存的條件。登上陸地後, 光合生物的進化速度大大加快, 在大約5億年內就從原始的陸地植物發展到高等的種子植物的全過程。
藻類植物的特徵
藻類植物體大小懸殊, 最小的直徑只有1~2微米,小到肉眼見不到, 而最大的長達60多米;
形態相差很大,有單細胞、群體和多細胞。群體各體由許多單細胞個體群集而成。多細胞個體有絲狀體、囊狀體和皮殼狀體等, 也有類似根、莖、葉的外形,
但不具備高等植物那樣的內部構造和功能。
藻類植物細胞含有各式各樣的色素,
而不同的色素組成標誌著進化的不同方向,是分門(phylum)的主要依據。但所有的藻類都含有葉綠素a和光合作用系統II並能利用水作為氫的供體,在光合作用中釋放出氧氣。現在大氣中的游離氧氣主要是光合作用的產物,其中大半是藻類所產生。藻類的色素主要有
4 類: 葉綠素、藻膽蛋白、胡蘿蔔素和葉黃素, 其中除葉綠素 a 以外, β-胡蘿蔔素也普遍存在於各種藻類, 只是在隱藻門數量較少而已。此外 ,
藍藻門、缸藻門和隱藻門還含有藻膽蛋白 , 隱藻門、甲藻門、黃藻門、金藻門、矽藻門和褐藻門含有葉綠素c, 原綠藻門、裸藻門、綠藻門和輪藻門含有葉綠素b,
在紅藻門有的種類則含有葉綠素 d 。少數藻類在演化 過程中營腐生或寄生生活 , 逐漸失掉葉綠素, 成為沒有色素的藻類。
藻類植物的生殖有營養體生殖、無性生殖和有性生殖。營養體生殖方法很多: 有特殊的營養枝, 如黑頂藻的繁殖枝, 掉地後則獨立生長為新的個體; 有依靠假子,
紅藻門的四分抱子,綠藻門的厚壁抱子等。有性生殖依靠配子,可以是同配或異配。同配由形狀大小一樣的配子相互接近, 融合形成厚壁的合子, 而異配則由大小不 同,
甚至形狀不一樣的配子融合形成合子。 卵配是一種異配, 其雌性細胞較大 , 一般不能游動, 而其雄性細胞較小, 有兩根鞭毛, 能自由遊動。紅藻的卵配尤其特殊,
卵囊稱果胞, 為一瓶狀構造, 卵在瓶底, 瓶頸即受精絲, 而精子在精子囊內, 不能游動,但可以隨水漂流,遇於受精絲則粘著上,
精子破囊而出,順著受精絲進入果胞與卵子結合成為合子, 後者立即發育成為一個雙倍體的果孢子體, 寄生在雌性 個體上。果孢子體成熟產生果孢子,
發育則成為獨立的孢子體。
一般分藻類植物為浮游藻類、飄浮藻類和底棲藻類。有的藻類, 如矽藻門、甲藻門和綠藻門的單細胞種類以及藍藻門的一些絲狀的種類浮游生長在海洋、江河和湖泊,
稱為浮游藻類。有的藻類如馬尾藻類飄浮生長在馬尾藻海上, 稱為飄浮藻類。有的藻類則固著生長在一定基質上稱為底棲藻類, 如藍藻門、紅藻門、
褐藻門、綠藻門的多數種類生長在海岸帶上;這些底棲藻類在一些地方形成了帶狀分佈, 一般說在潮間帶的上部為藍藻及綠藻,
中部為褐藻而下部則為紅藻。但中國海岸帶海域和亞熱帶海域的冬春兩季, 高潮帶常有藍藻門的鬚藻, 紅藻門的紫菜、小石花菜, 褐藻門的鼠尾藻, 綠藻門的綠苔、滸苔;
中潮帶常有紅藻門的海蘿, 褐藻門的萱藻和綠藻門的礁膜、石蒓等。低潮帶及潮下帶種類很多, 如紅藻門的石花菜、角叉藻、多管藻、凹頂藻 ,
褐藻門的海帶、裙帶菜、海蒿子和綠藻門的海松。潮間帶還有許多石沼, 為藻類的生長提供了良好的條件。還有兩種特殊的生態環境適宜於若干藻類群落的生長:
如亞熱帶和熱帶的 紅樹林, 常有卷枝藻、鏈藻、鷓鴣菜在氣根上及樹幹基部上生長, 熱帶海洋的珊珊礁常有大量的仙掌藻屬植物。
在淡水的瀑布, 滴水的岩礁以及急流的江河, 都可以找到多種藻類的生長, 有藍藻門、紅藻門和綠藻門的種類。 在緩流和靜水的環境如湖泊、水庫及積水處常可以找到網藻、鞘藻、水棉等綠藻。在溫泉水溫一般超過 60 ℃ , 可找到某些藍藻。有些藍藻還生長在沙漠及南北極砂礫 內部。土壤藻類有藍藻門、矽藻門和綠藻門各有 20 多種。在潮濕牆上、樹枝上、樹幹上也可以找到多種藻類。有的藻類還同菌類共生成為地衣。可以認為, 藻類植物是無所不在的。
眼蟲植物門 Phylum: Euglenophyta
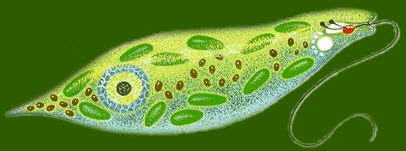
眼蟲(游藻)綱
Class: Euglenophyceae
眼蟲綱在其成年階段都有一根、幾根或許多根鞭毛作為其運動細胞器,
由於它們營養方式及生殖方式的原始性和多樣性,動物學上一般認為眼蟲(稱為鞭毛蟲綱 Mastigophora)是最原始的原生動物,
而且通過它可以將動物與植物界聯繫起來。
眼蟲具有色素體 , 可行光合作用即進行植物性營養。澱粉及副澱粉是其主要的食物貯存物, 有無性生殖和有性生殖 , 藻體通常具有2根鞭毛, 體表為皮膜或纖
維素細胞壁, 自由生活。
(1) 眼蟲目
Euglenales

綠眼蟲 (綠眼藻) Euglena viridis Ehrenb., 眼蟲科: Euglenaceae (館藏編號: NSF-320)
單細胞藻類,
全長20至300微米(μm), 廣布於淡水中。身體呈卵圓形、球形、長圓形等 , 通常能分出前後端
, 都以鞭毛作為運動器官 , 體表具細胞膜、或皮膜或細胞壁。體表具有皮膜 , 是由一層 蛋白質組成螺旋形排列的條紋, 在條紋上有晴和溝, 使皮膜呈現斜紋,
眼蟲的皮膜很薄、富有彈性 , 使身體可以收縮改變形狀, 做眼蟲式運動 (euglenoid movement) 。鞭毛蟲缺乏明顯的內質與外質的分化。
身體的前端有一凹陷的溝, 稱儲蓄泡 (reservoir) 或稱胞咽, 但它不是取食的器官, 是水的貯存處及鞭毛著生部位, 基體位於它的基部 ,
由基體伸出兩根鞭毛, 一根很長伸向體外, 一根很短不伸出儲蓄泡, 身體的前端含有類胡蘿蔔素 (carotinoid) 的脂類球集合而成的紅色眼點,
這種包素球只有在基質中含有很少的氮及磷時才出現, 否則將消失, 它可以吸收光線。在鞭毛的基部具有副鞭毛體
(paraflagelIar body), 是一種光敏感結構 , 當一側的光被眼點吸收後, 可使它產生一個定向的光反應,
所以副鞭毛體與眼點構成了眼蟲類的感光細胞器。
身體前端有一個或多個伸縮泡, 它是由幾個小泡收集了細胞質內過多的水分愈合而成, 當伸縮泡充滿水分之後便排到儲蓄泡中,
再通過其開口排到體外。細胞內散佈有形狀、數目不等的色素體, 色素體中含有葉綠素 a 和 b, 胡蘿蔔素 (carotin) 和葉黃素, 使蟲體表現出綠色。
具有一個大的細胞核, 位於身體的近後端, 核內有核內質( edosome)。眼蟲的細胞核在分裂間期時染色質濃縮, 分裂過程中不行成紡錘體, 核膜不會消失,
這是很原始的原生生物生殖現象。
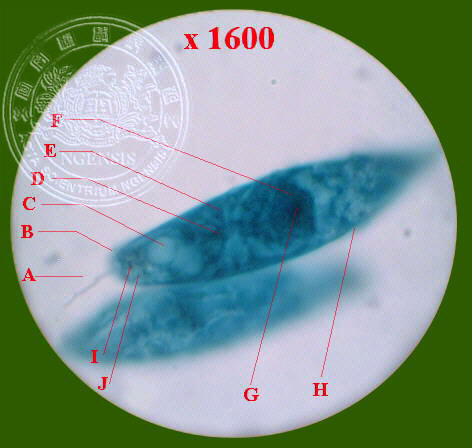
本室拍攝較為清晰的眼蟲光學顯微鏡結構的圖解 (22
VI NSC14) :
A.鞭毛
B.胞口 C.伸縮泡 D.副澱粉粒
E.葉綠素體 F.細胞核 G.核內質 H.皮膜
I.眼點 J.儲蓄泡
黃藻植物門 Phylum: Chrysophyta
黃綠藻綱
Class: Xanthophyceae
(1) 異管藻目 Heterosiphonales
無隔藻 Vancheria sp., 無隔藻科: Vaucheriaceae (館藏編號: NSF-323)
矽藻綱
Class: Bacillariophyceae (Diatoms)
(1) 圓心矽藻目
Centrales
盤狀矽藻亞目 Discinae

小環藻 Cyclotella sp., 盤狀矽藻藻科: Coscinodiscaceae (館藏編號: 未編號 )
(2) 羽紋矽藻目
Pennales
2a.無脊溝亞目
Araphidineae
扇矽藻 Licmophora sp., 帶矽藻科: Fragilariaceae (館藏編號: NSF-321)

脆杆藻 Fragillaria sp., 帶矽藻科: Fragilariaceae (館藏編號: cf. NSF-326 / 4)
2b.兩脊溝亞目 Biraphidineae
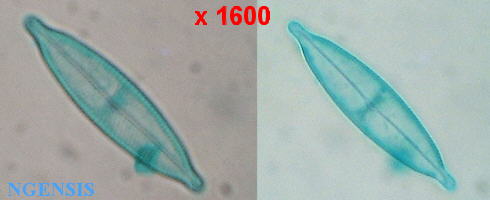
扇吻嘴羽紋矽藻 Navicula rhyncocephala Kutz, 羽紋矽藻科: Naviculaceae (館藏編號: cf. NSF-326 / 1)

舟形藻 Cymbella sp., 羽紋矽藻科: Naviculaceae (館藏編號: cf. NSF-326 / 2)

尼釆藻 Nitzschia sp., 尼釆藻科: Nitzschiaceae (館藏編號: cf. NSF-326 / 3)
綠藻植物門 Phylum: Chlorophyta
綠藻綱 Class: Chlorophyceae
(1) 團藻目
Volvocales
衣藻亞目
Chlamydomonadinae
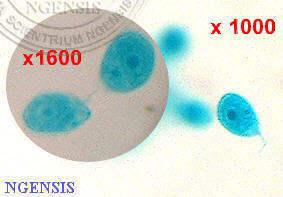
衣藻
(單胞藻)
Chlamydomonas
sp., 單胞藻科: Chlamydomonadaceae
(館藏編號:
NSF-322)
單細胞藻類,
動物學者亦稱之為衣滴蟲, 藉鞭毛在水中活動。很有趣的是: 從植物學的角度觀之, 它和眼蟲的親緣關係甚遠;
但動物學者則認為它們皆以鞭毛在水中活動及彼此的生態特徵很接近而將之併入鞭毛蟲綱中。
(2) 披髮藻目
Ulotrichales
石蒓亞目 Ulvales

條滸苔 Enteromorpha clathrata (Roth.) J. Agardh, 石蒓科: Ulvaceae (館藏編號: NSF-246)

扁滸苔 Enteromorpha compressa (L.) Greville, 石蒓科: Ulvaceae (館藏編號: NSF-260)

石蒓 Ulva lactuca L., 石蒓科: Ulvaceae (館藏編號: NSF-247)

裂片石蒓 Ulva fasciata Delile, 石蒓科: Ulvaceae (館藏編號: NSF-268)
(3) 剛毛藻目 Cladophrales

剛毛藻 Cladophora sp., 剛毛藻科: Cladophoraceae (館藏編號: NSF-324)
(4) 接合藻目
Conjugales
4a.
真接合藻亞目
Euconjugatae

水綿
Spirogyra jugalis,
接合藻科:
Zygnemataceae
(館藏編號:
NSF-325)
由許多細胞連結而成的絲狀體, 水綿的各個細胞中彼此可以獨立生活,
生殖時兩條水綿平時彼此並排,生殖時由其中一條中細胞的細胞質流入另一方的細胞中而形成接合子,之後便進入休眠狀態;接合子待到春天時就發芽。(右圖:
水綿單體中的細胞核)
4b.
鼓藻亞目 Desmidiineae
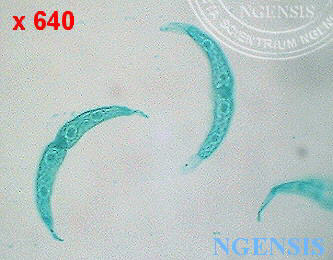
鼓藻 Cosmarium sp., 鼓藻科: Desmidaceae (館藏編號: NSF-326)
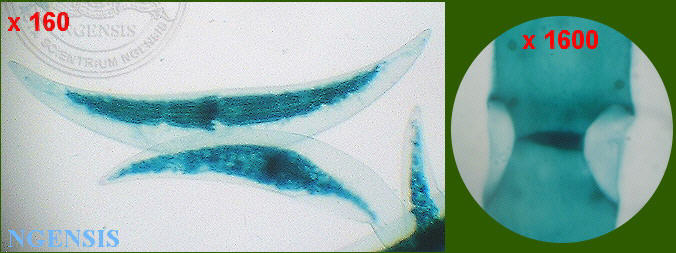
新月藻
Closterium
sp., 鼓藻科:
Desmidaceae
(館藏編號:
NSF-327)
新月藻雖然是單細胞生物, 但生殖時可以由兩個個體接合成接合子。接合開始時首由先兩個細胞相向並排,之後各細胞壁由中間裂開
(如右圖),其中的細胞質即時進行交換而合成一體。結合後細胞變成圓形,
一層厚壁亦隨即生成形成接合子。
(4) 管藻目 Siphonales
水松 Codium fragile Hariot, 水松科 Codiaceae (館藏編號: 暫未編號 )
褐藻植物門Phylum: Phaeophyta
(Melanophyceae)
同型世代綱 Class : Isogeneratae
(1) 外子藻目 Ectocarpales

紅水雲 Ectocarpus rhodochortonoides Boergesen, 水雲科: Ectocarpaceae (館藏編號: NSF-273)
(2) 網目藻目 Dictyotales

裸胞藻 Pocockiella variegata (Lamouroux) Papenfuss, 網目藻科: Dictyotaceae (館藏編號: NSF-262)

大團扇藻 Padina crassa Yamada, 網目藻科: Dictyotaceae (館藏編號: NSF-263)
(3) 石衣藻目 Fucales
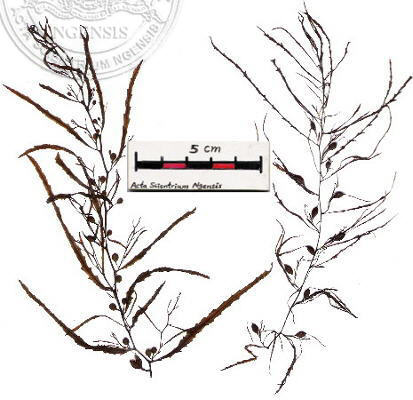
狹葉馬尾藻 Sargassum angustifolium (Turn.) C. Agardh, 馬尾藻科: Sargassaceae (館藏編號: NSF-245)
紅藻植物門 Phylum: Rhodophyta
原始紅藻綱 Class: Protoflorideophyceae (Protoflorideae)
(1) 頭髮菜目
Bangiales

長葉紫菜 Porphyra dentata Kjellman, 頭髮菜科: Bangiaceae (館藏編號: NSF-270)

圓葉紫菜 Porphyra suborbiculata Kjellman, 頭髮菜科: Bangiaceae (館藏編號: NSF-271)
真紅藻綱 Class: Florideophyceae (Florideae)
(1) 海索麵目 Nemalionales

粗狀頂毛藻 Acrochaetium robustum Boergesen, 頂毛藻科: Acrochaetiaceae (館藏編號: NSF-264)
(2) 隱絲藻目 Cryptonemiales

花布糊 Gloiopeltis complanata Yam, 剌枝藻科 Endocladiaceae (館藏編號: 暫未編號 )
珊瑚藻 Corallina pilulifera, 珊瑚藻科 Corallinaceae (館藏編號: 暫未編號 )
(3) 杉海苔目 Gigartinales
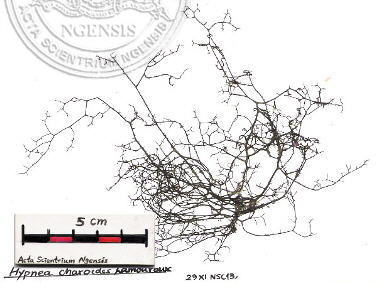
長枝棘糊藻 Hypnea charoides Lamouroux, 棘糊藻科: Hynpeaceae (館藏編號: NSF-261)
(3) 仙菜目
Ceramiales

縱胞藻 Centroceras clavulatum (C.Ag) Mont., 仙菜科: Ceramiaceae (館藏編號: NSF-258)

小仙菜 Ceramium byssoideum Harvey, 仙菜科: Ceramiaceae (館藏編號: NSF-269)
輪藻植物門 Phylum: Charophyta
輪藻植物綱 Class: Charophyceae
(1) 輪藻目 Charales

麗 藻
Nitella sp., 輪藻科: Characeae (館藏編號:
NSF-954)
按
Hong Kong Freshwater Plants (p.27, 1978)
所載這種植物為 Chara spp.,
根據所採與有關圖片極為相似物種的顯微鏡檢察得知, 有關物種不是輪藻屬( Chara )而是麗藻。
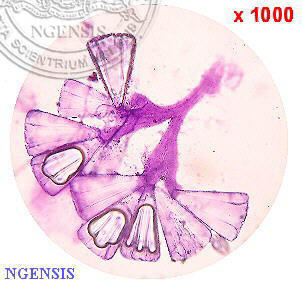
圖片說明
A. 標本(部分); B.植株之一部分;
C.成熟枝之一部分; D、E.
短枝的一部分;
F. 卵囊
主要參考文獻
[1]
Strasburger's Textbook of botany,
rewritten by Dietrich von Denffer ... [et al.] ; transl. from the thirtieth
German ed. by Peter Bell, David Coombe, Fischer ; London
: Longman Group Limited, 1976.
[2]《中國大百科全書》生物學
I, II, III,
中國大百科全書出版社,北京.上海,1991.12.
[3]《簡明生物學辭典》吳熙載 主編,
湖北人民出版社,
1985/01.
[4]《植物分類學
地衣及藻類》卷貳, 劉棠瑞 著, 國立編譯館, 中華民國七十四年十二月初版
[5]《海洋生物 ~ 藻類》張義浩等主編,
淅江大學出版社, 11/2002.

本網頁中所有圖片及文本受延陵科學綜合室版權保障
All images and text on this page are
copyright protected © Acta Scientrium Ngensis